The Human Brain
Cerebellum and Basal Ganglia
Structure of the Cerebellum

The cerebellum (red) is a solid structure attached to the back of the brainstem by two cerebellar peduncles - one on each side of the midline. These contain the axons by which the cerebellum receives information, and passes information back to the brainstem and cerebral hemispheres.

The cerebellum is divided into three lobes:
- The Flocculonodular lobe - the oldest in evolutionary terms, and the most caudal, sometimes called the vestibulo-cerebellum, because of its association with the vestibular system
- The Anterior lobe, sometimes called the spino-cerebellum, because of its association with spinal inputs related to proprioception
- The Posterior lobe - the most recent development, concerned with inputs from the cortex, particularly with visual, auditory and somatosensory inputs related with body position and movement.
In the midline is the vermis (the nodule in the flocculonodular lobe). and laterally are the hemispheres; the area on each side of the vermis is sometimes called the intermediate lobe
| Evolution of the Cerebellum |
|---|
The oldest part of the cerebellum is the flocculonodular lobe, which has connections with the vestibular apparatus and is particularly concerned with balance and eye movements. |
The anterior lobe has connections with the spinal cord and is concerned with coordination of the musculature. |
The posterior lobe developed greatly when primates evolved and adopted the upright posture; at the same time the forebrain expanded and there are important connections between these two. |
The cerebellum is concerned with the control of movement, integrating signals from different parts of the nervous system and generating error signals that allow adjustments to be made to achieve the desired objective of the movement. |
In cerebellar disease, voluntary movements can become grossly exaggereated and unsuccessful attempts to correct them result in a tremor that occurs during voluntary movement. |
| Divisions and Nuclei of the Cerebellum |
|---|
| 1.
The large lateral hemispheres of the posterior lobe project to the dentate nuclei, and onwards to the cerebral cortex, via the red nucleus of the midbrain 
Flocculo-nodular lobe (vesibulocerebellum) projects to vestibular areas of the brainstem via the fastigial nuclei. The interposed nuclei (N interpositus- emboliform and globose nuclei) are the output pathway for the intermediate area of cerebellar cortex, lateral to the vermis. |
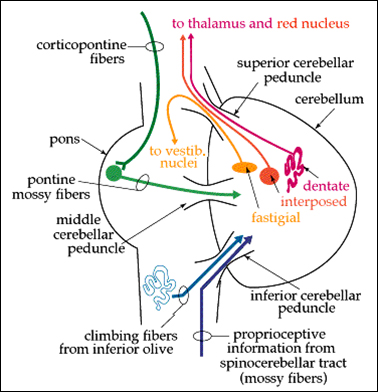
| Connections between the Cortex and Cerebellum |
|---|
|
The cerebellum receives copies of the messages (motor commands) sent down the corticospinal tract. |
|
The cerebellum compares these commands, with feedback information (information from proprioceptors, vision, touch, hearing) and decides if the command is being accomplished, and whether there are errors. |
|
Information about any errors is returned to the cortex, so that it can adjust the command signal. |
|
The first pathway - a copy of the command message issued by the motor cortex - involves cortico-pontine and ponto-cerebellar neurones. |
|
The sifting of feedback signals to extract information about errors is done by Purkinje cells, the principal cell of each cerebellar module. The pontine fibres that feed into the cerebellum are known as 'mossy fibres' because of the ultrastructure of their terminals. Mossy fibres supply the cerebellar cortex with information from many areas of the cortex, concerned with the position of the body, information from the visual and auditory fields, and the position of objects that are touching the skin. |
|
A second input to the cerebellum arises in the inferior olive (in the medulla) and these fibres climb around the Purkinje cells and are known as 'climbing fibres'. The two inputs to the cerebellum are compared within the cerebellar cortex, so as to provide the cortex with information about errors in movement. The climbing fibres may also be concerned with selecting appropriate movements while learning motor skills. |
|
The communication of error signals to the cortex depends on the output of Purkinje cells, through the deep cerebellar nuclei to the cortex (with synapses in the red nucleus and the thalamus). |

| The Cerebellar Cortex |
|---|
The cerebellar cortex has three layers -
|
The Cerebellar Cortex is where comparisons are made between the command signal and the feedback signals from proprioceptors, touch, vision and hearing. |
Granule cells in the cerebellar cortex have axons that pass towards the surface of the molecular layer and bifurcate, sending their collaterals in opposite directions within the molecular layer; there are enormous numbers of parallel fibres in the molecular layer. |




